
Aequorea victoria jellyfish (image courtesy of http://home.sandiego.edu/~cloer/nobel08.html)
Since the isolation of the first naturally fluorescing protein from jelly fish in the 1960s, fluorescent proteins (FPs) have become integral to a wide range of biological/medical applications, allowing visualization of biomolecules and processes in living cells and tissues to a much greater degree than was previously possible. Extensive molecular engineering of naturally occurring FPs has led to several families of recombinant proteins tailored to specific biophysical and imaging purposes. Fine-tuning of the activation and multimerization properties of FPs has helped to alleviate some issues of “photo-bleaching” (loss of light emission over time) and cellular toxicity. FPs that emit light of a wide range of different wavelengths have been isolated or engineered. In particular, FPs that fluoresce in the red to far-red range are of interest because they can be used in conjunction with existing, well-characterized green, blue, or yellow emitting FPs in co-labeling experiments. In addition, most cells naturally contain molecules that auto-fluoresce in the green and blue range and red light better penetrates animal tissues for deep imaging.
Our group is particularly interested in whether FPs can be engineered to have photophysical properties, such as brightness, that are superior to those of conventional organic fluorophores, and if so, what are the structural and dynamical features of FPs that enable these properties? We use a combination of random and targeted mutagenesis to create and assess new red/far-red FPs with improved spectral and cellular properties. We have designed and operated several novel microfluidic systems for screening these large libraries of mutants. For example, microdroplets containing 1–3 bacterial cells, each expressing a unique protein variant, can be individually excited by a chosen laser wavelength, their emission assessed via a customized detection system, and the droplets/cells of interest collected through a gated channel. Thus we can enhance for a cell population carrying FPs with particular qualities.


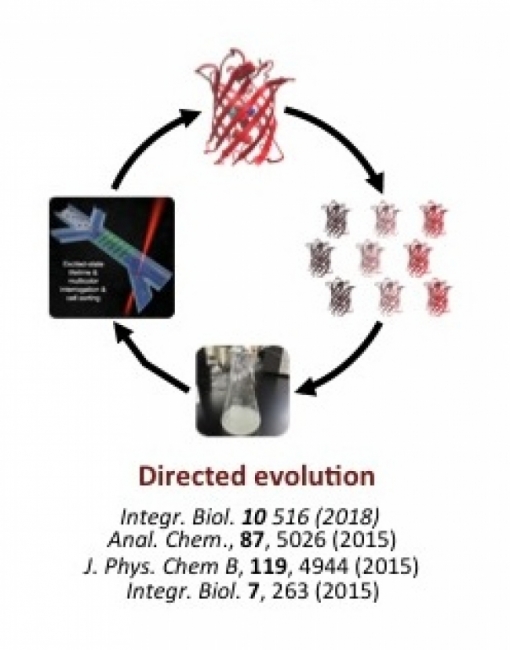


Similarly, we have designed and fabricated microwell devices that allow excitation and emission analysis of droplets containing several million molecules of purified FP. This unique microarray system has been designed to operate on spatial and time scales that allow us to examine factors that influence the maturation/activation of the fluorescing region of the protein complex, the chromophore, and the proteins tendency to turn “off” (reversibly or reversibly convert to a dark state). This information will inform further protein-engineering strategies. Conversely, we are interested in exploiting the “switchable” quality of fluorophores to create biosensors to detect specific analytes or processes in living cells.
We have applied these techniques to identifying key amino acids that contribute to the stability and activity of multiple red FPs. We can tweak our assays to select for mutants with desired spectral qualities, such as increased fluorescence lifetime, increased fluorescence emission (radiative rate) or, conversely, decrease in the energy lost through non-fluorescing processes (non-radiative rate). We also examine mutant proteins that show promise in vitro in living cells (bacteria, yeast and human) to further select for desirable behaviors, such as increased overall or subcellular brightness, better chromophore maturation kinetics or decreased toxicity.